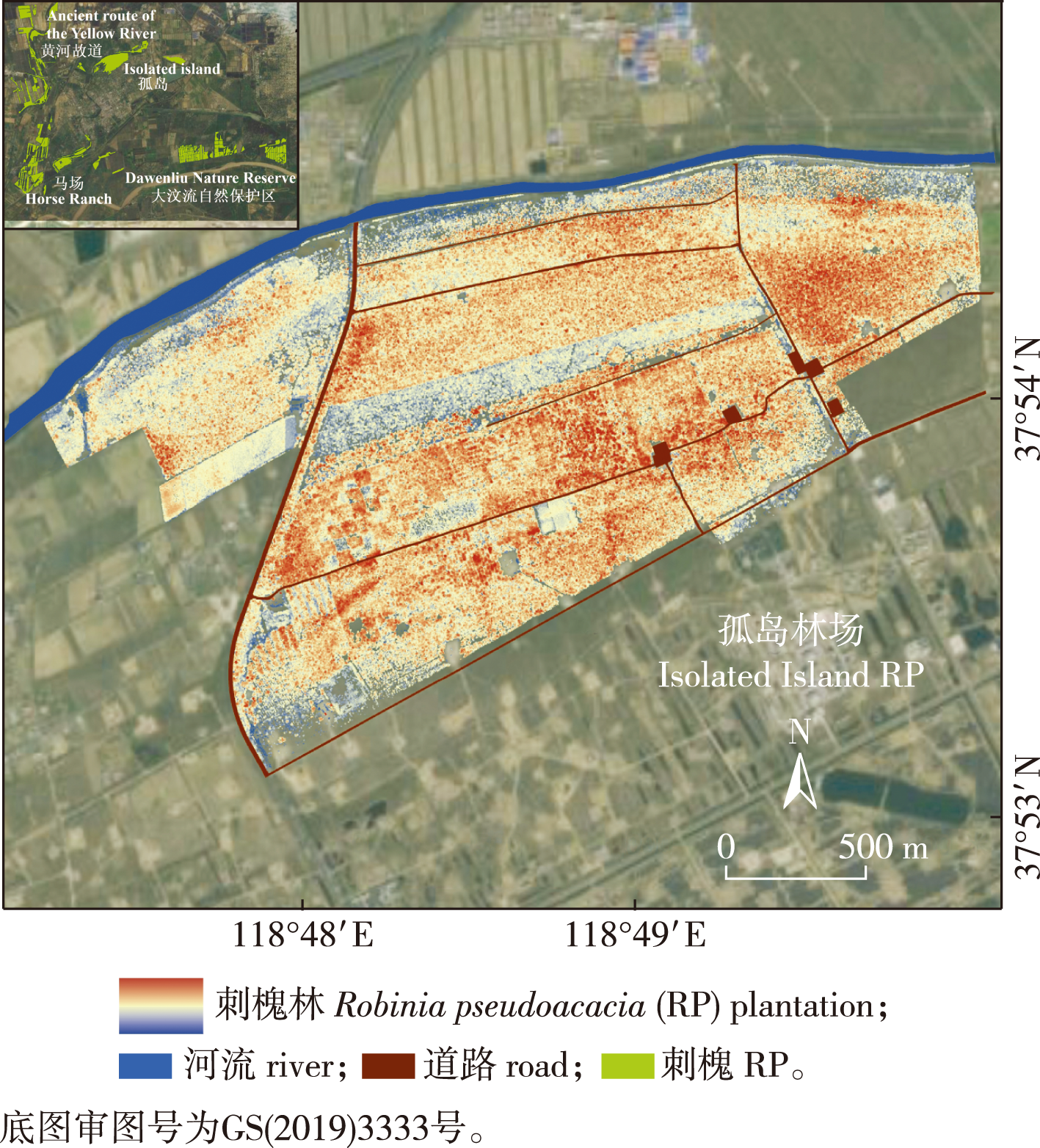
【目的】边缘效应是指在不同生态系统交界处,由于生态因子的差异及相互作用,引发系统内某些组分显著变化的现象。这一效应广泛存在于森林、湿地、草原等生态系统中。研究森林边缘效应对生态保护、营林管理、气候变化应对和碳储量计算具有重要意义。【方法】以黄河三角洲刺槐人工林为研究对象,利用UAV-LiDAR和Sentinel-2遥感数据,分别提取7个结构指标和3个生理指标,量化分析人工林三维结构和生态功能的边缘效应,并探讨不同边缘类型的边缘效应特点。【结果】①边缘效应对刺槐人工林分结构产生了负面影响。边缘区域的林分树高降低1~5 m、覆盖度减少7%~53%、叶面积指数(LAI)下降10%~77%,林分密度下降6%~52%,同时其冠幅面积和体积也减小。②边缘效应对森林生态功能也产生了负面影响,如削弱了植被的光合作用,其吸收的光合有效辐射(FAPAR)降低8%~50%,叶绿素含量下降9%~72%,冠层含水量也下降8%~50%。③不同边缘类型的影响范围不一样,河流边缘的影响范围可达150 m,而道路边缘影响强度较小,范围约为30 m。【结论】边缘效应对黄河三角洲滨海防护林的植被结构和生态系统功能产生了多维度影响。本研究有助于滨海防护林管理和生态保护,帮助应对气候变化与区域环境退化。
【Objective】The edge effect refers to the phenomenon that some components within the ecosystems change significantly due to the differences and interactions of ecological factors at the junction of different ecosystems. This phenomenon is ubiquitously observed across diverse ecosystems including forests, wetlands and grasslands. The investigation of forest edge effects holds substantial importance for advancing ecological conservation efforts, enhancing forestry management practices, addressing climate change impacts, and refining carbon storage estimations.【Method】This study quantified edge effects in Robinia pseudoacacia plantations in the Yellow River delta using UAV-LiDAR and Sentinel-2 data. Seven structural and three physiological indices were analyzed to assess 3D structural and ecological functional variations, with distinct edge effect patterns identified across different edge types.【Result】(1) The edge effect significantly altered vegetation structural parameters, with observed reductions of 1-5 m in tree height, 7%-53% in vegetation coverage, 10%-77% in leaf area index (LAI), and 6%-52% in tree density. Canopy structural features, including projected area and volume, also exhibited marked decreases. (2) Ecological functions were substantially compromised at forest edges, manifested by impaired photosynthetic capacity with 8%-50% reduction in fraction of absorbed photosynthetically active radiation (FAPAR), 9%-72% decline in chlorophyll content, and 8%-50% decrease in canopy water content. (3) Edge influence zones showed distinct spatial patterns, with river edges demonstrating an extensive impact range of 150 meters, contrasting with the relatively limited 30-meter influence zone of road edges.【Conclusion】The edge effect exerts multidimensional influences on both structural characteristics and ecosystem functionalities of coastal shelterbelts in the Yellow River Delta. This research provides critical insights for enhancing coastal shelterbelt management and conservation strategies, facilitating the optimization of sustainable forest plantation development, and contributing to climate change mitigation and regional environmental restoration efforts.
 PDF(4043 KB)
PDF(4043 KB)